魚類の鱗は真皮の内部に発達した骨格=皮骨であり
遂にハリセンボンの鱗は棘に変形した。
魚の鱗は相互に動く鱗は重く厚い鱗から軽く薄い鱗へと進化して、
面的なテンセグリティ構造を形成する。
水の抵抗を減衰させ迅速に移動し
水圧に耐えながら敵の攻撃から自身を守るために。
鱗は真皮が変化した骨格=皮骨


魚類の鱗は真皮の内部に発達した骨格=皮骨であり
遂にハリセンボンの鱗は棘に変形した。
魚の鱗は相互に動く鱗は重く厚い鱗から軽く薄い鱗へと進化して、
面的なテンセグリティ構造を形成する。
水の抵抗を減衰させ迅速に移動し
水圧に耐えながら敵の攻撃から自身を守るために。
鱗は真皮が変化した骨格=皮骨
フグの4面体状モジュールは自然のテンセグリティ・デザインである。
バックミンスター・フラーは、8面体状のテンセグリティモジュールからなる
球状テンセグリティ構造(1982年)をデザインしている。
約40年前私はカリフォルニアでこのモデルの修復に従事したことがある。

無脊椎動物や藻類などを食べるフグは脊椎動物であるが、
驚くことに連続した脊椎をほとんど形成していない。
すべてのテトラ状の骨は張力材ネットワーク上に
浮かんでいることになる。
テンセグリティ構造は大黒柱のない分散型の無脊椎構造である。

tensegrity
伸縮しやすい皮膚を採用したフグ(Blowfish)は、
伸縮自在の胃に素早く大量の水や空気を吸入させて
浮遊するテンセグリティ構造を採用している

フグ(Blowfish)の多数の棘はテトラ状の骨であり
その骨は互いに非接触な圧縮材を形成する。
フグの筋肉と皮膚はそれらを統合する連続した張力材となる。
フグは収縮拡大するテンセグリティ構造を採用している。
この構造は海表面でも安全に浮遊することができる。
不連続なテトラ状の骨モジュール
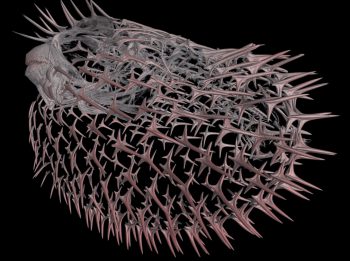
非対称性が均衡を破壊しているように見えるのは
あるいは、対称性が破れているように見えるのは
観察者の局所的な経験によって制限されている可能性がある。
シナジェティクスは、
反対称性と非対称性(鏡像対称性)とを明確に分離できる
動力学的なシナジェティクスモデルを1981年に発見している。
反対称性の動くシナジェティクスモデル
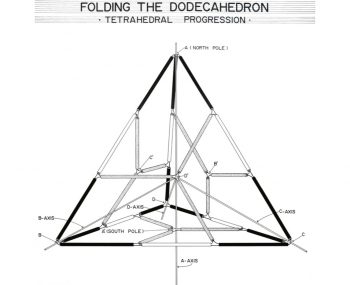
反対称性のシナジェティクスモデルが発見されも
それまでの対称性の概念がまだ生き残っているのは
社会的な認識上のタイムラグではなく
モデルによる思考言語の破壊の方法にある。
反対称性のシナジェティクスモデル
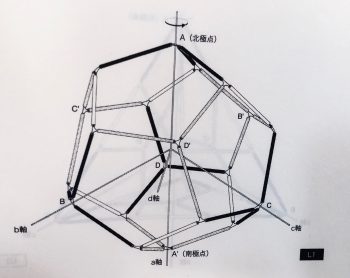
正12面体状の準結晶構造は安定した構造である。
その構造の特性を証明する空間充填モジュールを科学者はまだ発見していない。
正12面体状の準結晶構造

準結晶の構造では周期性と非周期性は共存する。
シナジェティクスは、「準結晶構造は五回対称性を有する
オクテットトラスの階層構造である」
という数学的証明をシナジェティクスモジュールによって達成した。
科学者は正12面体状の準結晶構造を形成する
空間充填モジュールを発見できなかった。
準結晶構造 異なる3種の金属元素から形成されるトラス構造

遅延は頭脳メカニズムの限界から生まれる。
遅延は無の間隔であり逸脱に属する。
現象を認識するには、
残像の生成とその再生を待たなくてはならない。
シナジェティクスモデルにはモデル現象学がある。
多頂点体の「対称性の破れ」の実験に私は5年間を要した。
多頂点体の「対称性の破れ」の実験 1989 梶川泰司 シナジェティクス研究所